Research
Vertebrate Paleontology & Evolution
My research is primarily focused on exploring the diversity and distribution of birds throughout the Cenozoic Era. This comprises anatomical studies and descriptions of avian fossils, and inclusion of both extinct and extant birds in combined phylogenetic analyses of morphological and molecular sequence data. Complementary lines of inquiry include issues related to using fossils as calibrations for divergence time analyses (i.e., dating phylogenetic trees) and comparisons of the sensory systems and bone microstructure of living and extinct birds.
Avian Paleobiology
The last thirty years have been marked by an astounding increase in our knowledge of the clade Aves and in the nuance with which that resulting wealth of information is interpreted. New fossil specimens that provide insight into the species diversity and biogeographical history of birds are constantly being discovered and make the fossil record of birds (i.e., avian dinosaurs) an exciting area of current research. Although I have studied the fossil record and anatomy of several clades of birds, I have a particular interest in the fossil record and evolution of seabirds and shorebirds (go to publications). Because seabirds such as the wing-propelled diving Pan-Alcidae inhabit both sea and land during different parts of their life cycles, they contribute in unique ways to our understanding of the dynamic interactions between physical and biological Earth systems. For example, questions such as how climate warming (both past and present) and resulting higher sea levels affect coastally nesting seabird species are best addressed by the study of species that inhabit the boundary between land and sea biomes (i.e., shoreline environments). Subsequently, changes in extant avian species diversity, population sizes and distributions are best contextualized with knowledge from the paleoclimatic and fossil records.

The holotype specimen of the extinct, flightless, pelagic, wing-propelled diving seabird Mancalla vegrandis, a new species I described in 2011. (Smith, 2011, figures 7, 8).
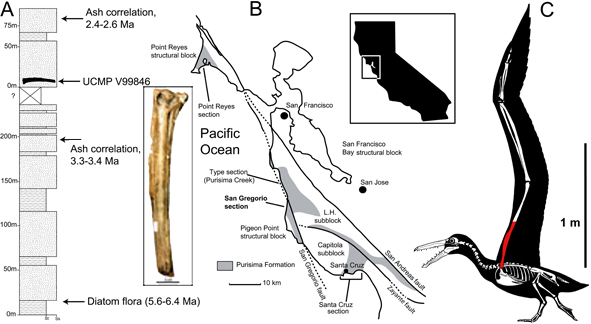
The Pelagornis fossil pictured here (see inset of UCMP V99846) represents a bird with the third largest wingspan currenty known. A colleague and I described the humerus of this giant pseudotoothed bird in 2011. Also pictured are the Stratagraphic horizon (A), geographic location (B) and a reconstruction of the bird (C; Boessenecker & Smith, 2011, figure 1; artwork by R. Boessenecker; reconstruction modified from that of C. Anzures in Mayr & Rubilar-Rogers, 2010).
Phylogenetic Systematics
Even though birds are arguably the most intensely studied group of organisms on Earth, relationships of birds remain a compelling area of new research and debate among systematists such as myself. Molecular sequence based methods have begun to resolve many longstanding issues of contention in avian systematics; however, the majority of extinct species have yet to be integrated into hypotheses of avian relationships. Just as knowledge of extant birds is critical to interpretations of behavior and other ecological variables in extinct organisms, the millions of years of evolutionary history documented in the fossil record facilitates research into broad evolutionary questions that can often not be addressed through studies that are limited to living organisms.
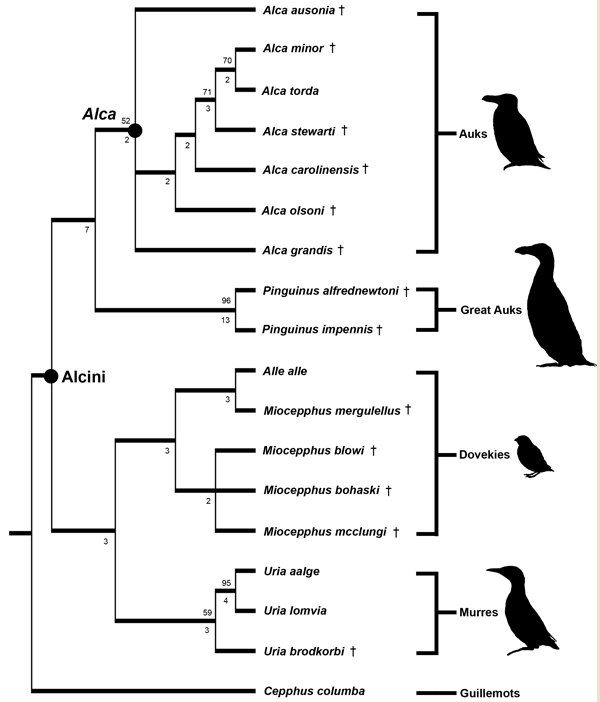
Phylogenetic hypothesis of relationships among extant and extinct Alcini (Charadriiformes, Pan-Alcidae; Smith & Clarke 2011, figure 6). Note that more than half of the species diversity in this clade is represented by extinct taxa (denoted by a "†")
Divergence Time Estimation Methods
Estimating the timing of major cladogenetic and biogeographic events through divergence time estimation analyses is one of the most ascendant yet controversial lines of inquiry in systematic biology. As part of my postdoctoral research at The National Evolutionary Synthesis Center (NESCent) I focused on identifying potential bias introduced by the temporal distribution of fossil calibrations in divergence analyses (read more here). Although previous work has alluded to this potential source of inaccuracy, detailed evaluation of this issue remains an area with significant promise to produce key new insights into the tempo of evolution, and refinement of divergence estimation methods. Additionally, quantification of potential bias created by the temporal distribution of fossil calibrations will lead to a more nuanced understanding of rate heterogeneity, an issue that remains at the core of developing more accurate methods of divergence estimation. Most importantly, divergence estimation facilitates comparisons between paloeclimatic events and evolutionary events. Because choice of fossils used as calibrations has been shown to be a significant source of potential introduced error in divergence time analyses, myself and a Divergence Dating Working Group of more than 20 other paleontologists, molecular biologists and computer programmers developed an online Fossil Calibration Database and associated scientific peer-reviewed journal (Palaeontologia Electronica - Fossil Calibration Series) that provides a venue to publish descriptions of new fossil calibrations.
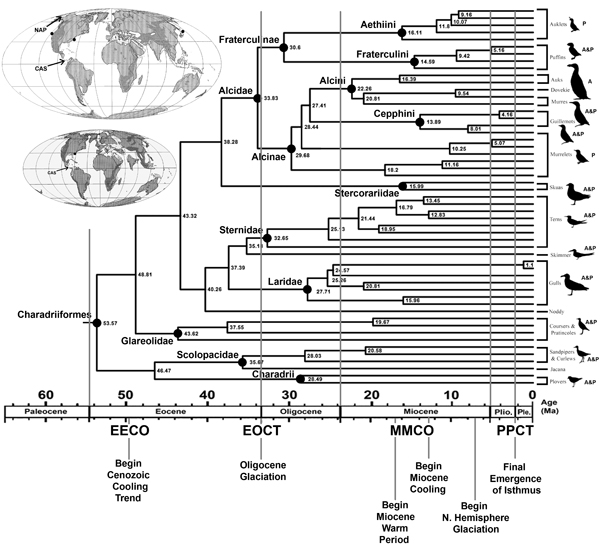
An example of a time calibrated tree for Charadriiformes and comparison with major paleoclimatic events. Abbreviations: (EECO) Early Eocene Climatic Optimum; (EOCT) Eocene-Oligocene Climatic Transition; (MMCO) Middle Miocene Climatic Optimum; (PPCT) Pliocene-Pleistocene Climate Transition. Silhouettes representing charadriiform clades are labeled according to extant geographic distribution: (A) Atlantic; (P) Pacific. Upper map shows Middle Miocene (~10 Ma) continental reconstruction with Northern Atlantic Passage (NAP) and Central American Seaway (CAS) both open. Miocene alcid fossils have been found on the eastern and western coasts of North America and Japan (black dots). Lower map shows Late Eocene continental reconstruction with Northern Atlantic Passage (NAP) closed and the sole Eocene alcid fossil locality (Smith, 2011, figure 8.16).
Avian Neuroanatomy and Histology
Through the use of computed tomography (CT) scanning, fine morphological details of soft endocranial tissues of both extant and extinct taxa can be studied through construction of digital endocasts. Likewise, the microstructural characteristics of bones from living and extinct taxa can be compared by preparing histological thin-sections of osseous tissues to be examined with a microscope. These methods allow me to discern anatomical details that might otherwise be difficult or impossible to document using traditional dissection techniques and facilitate the discovery of correlations between anatomy, ethology, and ecological traits.

Endocranial endocasts of the Razorbill Auk Alca torda, Great Auk Pinguinus impennis, Black Skimmer Rynchops niger, and Killdeer Charadrius vociferus in right lateral view (not to scale for comparison). These four endocasts illustrate the sensory system morphological differences between aquatic versus terrestrially foraging charadriiforms, between R. niger and all other charadriiforms, between wing-propelled diving pan-alcids and other charadriiforms, and between volant and flightless pan-alcids (Smith & Clarke 2012, figure 15).

Photomicrograph of a histological thin-section from the femur of the extant Razorbill Auk Alca torda (Smith and Clarke, 2014).
Phylogenetic Comparative Methods
Because species do not represent independent data points in statistical analyses, comparisons of life history traits such as body mass and skeletal measurements are informed by phylogenetic hypotheses. A phylogenetic generalized least squares model (PGLS) is often used to explore the relationship between continuous variables (e.g., brain volume versus body mass) in the context a phylogeny derived from discrete characters (e.g., osteological morphology and/or molecular sequence data).
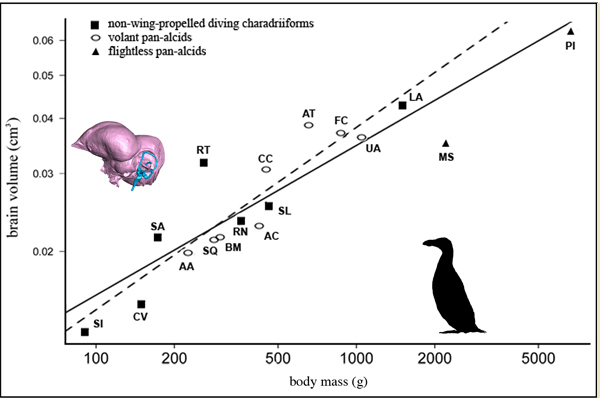
This PGLS scatterplot from Smith & Clarke (2012, figure 12) shows the different regression lines that are inferred when the relative body mass and brain volume of flightless and volant species are considered simultaneously (solid black line) and when volant species are considered independently (dashed line).